
Архозавры: Этозавры (Aetosauria).
Характеристика группы.
В позднем триасе архозавры стали не только доминирующими хищниками, но заняли несколько ниш растительноядных, которые до этого принадлежали дицинодонтам. Произошла смена фаунистических суперкомплексов на суше, и одно из главных мест в новом комплексе заняли Этозавры.
В целом, Этозавры (Aetosauria) – отряд круротарзальных архозавров и единственное его надсемейство Stagonolepididae. Этозавры появляются в начале позднего триаса (в Карнии), досигают рацвета в Нории, постепенно теряют многообразие в к кону Нория, становятся малочисленны в Раэтии и окончательно исчезают при переходе к Юрскому периоду. Но во времена своего расцвета этозавры были разнообразны и по размерам, и по способам питания. Среди них были и крупные травоядные, и более мелкие роющие формы, а некоторые могли приспособиться к питанию водной растительностью. Но не смотря на различия, все этозавры были панцирными рептилиями, покрытыми от головы до конца хвоста костяной броней.
Теперь подробнее посмотрим на этозавров. Очень интересен череп этозавров, который совершенно непохож на типичные черепа архозавров. Во-первых, начинается этот череп с какого-то пяточка – расширения предчелюстной кости, после которого морда вновь сужается. Считается, что пяточок мог быть адаптацией к подкапыванию корней растений. Во-вторых, для этозавров характерны очень большие ноздри, вытянутые в передне-заднем направлении. Такие ноздри могут быть дополнительной адаптацией к рытью, если ноздри зажимаются клапанами. Также они могут быть просто облегчением передней части черепа. В-третьих, у этозавров уменьшено предорбитальное окно и смещено вниз, так что слезная кость удлиняется и формирует верхнюю границу этого окна. В-четвертых, глазница круглая по форме, что нехарактерно для псевдозухий, имеющих чаще вытянутую сверху вниз глазницу. В-пятых, позади глазницы скула более или менее резко уходит вниз, иногда под углом близким к 45 градусам. В результате, челюстной сустав находится ниже конца зубного ряда. Чем-то это напоминает пермских диноцефалов. В-шестых, нижнее височное окно практически исчезает, от него остается маленький треугольный рудимент; из-за этого посторбитальная кость контактирует с квадратно-скуловой. В-седьмых, крыша черепа становится широкой и верхние височные окна становятся боковыми. В-восьмых, в нижней челюсти находится большое окно, вклинивающееся в зубную кость, однако в передней части данное окно не сквозное.
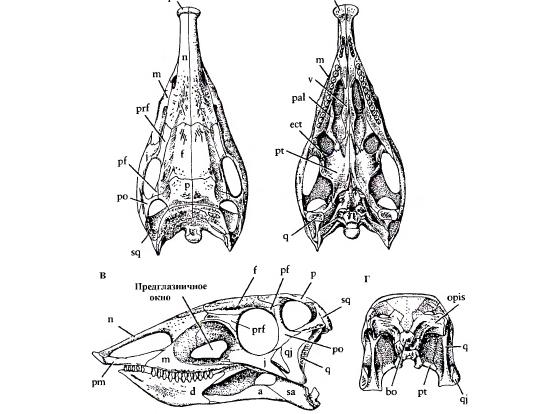
Рисунок черепа Стагонолеписа (Stagonolepis robertsoni). В этом типовом черепе обратите внимание на пятачок на конце морды, на отсутствие зубов, на огромные ноздри и уменьшившиеся боковые окна в черепе.
---
Этозавры потеряли большую часть зубов. На предчелюстной кости их нет, и на нижней челюсти зачастую нет зубов в передней части. Предполагается, что эти зубы могли быть заменены роговым клювом. Оставшиеся зубы в верхней и нижней челюсти стали мелкими и листовидными, с простой конической коронкой. Этозавры утратили текодонтность и их зубы прикреплены непостредственно к кромке челюсти. Такие зубы являются признаком питания мягкой растительностью, но, возможно, некоторые их обладатели питались мягкими беспозоночными.
ПОзвоночный столб этозавров особо ничем ни примечателен по сравнению с другими псевдозухиями, если не считать сокращения длины спинных отростков позвонков (из-за наличия брони) и удлинения боковых. В результате шея и хвост этозавров выглядели сплющенными.
Более интересны конечности этозавров. Что передние конечно. что задние очень мощные. По строению костей предполагается, что мышцы конечностей были сильно развиты. При этом предплечие у этозавров относительно укорочено по сравнению с плечом (как у современных роющих животных), а на плече выражены места прикрепления мышц, участвующих в копательных движениях. Однако, на 100% утверждать, что перечисленные выше адаптации связаны с рытьем не стоит, поскольку и голени у этозавров укорочены по сравнению с бедром. Возможно, это просто особенности строения, связанные с конструкцией панциря.
Задние конечности этозавров соединяют в себе прогрессивные и примитивные черты. Во-первых, таз у них был пластинчатым, примерно напоминал по форме таз эритрозухий. Но при этом передний край лобковой кости был усилен и впереди у нее был отросток, резко отогнутый вниз, как у попозавроидов. Мы не можем утверждать, что пластинчатое строение таза было архаичной чертой строения этозавров, возможно они вторично приобрели пластинчатый таз из-за особенностей своего строения. Во-вторых, стопа у этозавров имеет выраженный пяточный выступ и близка к симметричности; но при этом стопа вытянута за счет пальцев, и 5-я предплюсневая кость имеет выраженную крючковидность.

Остеодерма Lucasuchus hunti -- фотография. Типичная по форме спинная остеодерма этозавра. Обращаем внимание на центральный шип, который мог быть дополнительно покрыт роговым чехлом и на скульптирование остеодермы радиальными канавками и гребнями, расходящимися от шипа.
---
Но самое интересное в этозаврах – это их панцирь. Кроме двух рядов крупных пластин вдоль позвоночника этозавры получили боковые защитные пластины, брюшные защитные пластины и даже пластины, защищавшие снизу хвост. Спинные пластины были из них самыми крупными, они защищали до 80% спины животного, зато боковые пластины могли нести на себе более или менее развитые шипы. У некоторых представителей семейства Desmatosuchinae шипы в области шеи были развиты очень сильно и превращались в рога – приспособления активной защиты. костяные пластинки панциря были орнаментированы. Вид орнамента меняется от одного представителя этозавров к другому и служит систематическим признаком. В орнаменте встречаются локальные ямки, радиально расхрдящиеся гредни и канавки, а также центральные гребни и поднятия. Мы не знаем были ли покрыты остеодермы дополнительной роговой броней, но это вполне вероятно. Поскольку роговой чехол делал бы панцирь крепче, а шипы панципя были бы острее. Для укрепления панциря существовало не только налегание передних пластн на задние, но и частичное налегание боговых платин на спинные в области заднего конца спинных пластин.
В общем, этозавры являлись панцирными травоядными, занимавшими освобождающиеся исчезающими дицинодонтами экологические ниши. Защитный панцирь помогал им бороться с хищниками, но только самые крупные из этозавров могли всецело положиться на него. Они обзавелись огромными желудками (об этом мы судим по поперечным размерам крупных этозавром), помогавшими эффективно переваривать растительную пищу. Эти этозавры гарантированно передвигались на четырех конечностях. А вот более мелкие этозавры вполне могли передвигаться и на двух конечностях. Хотя типичным была квадрупедальность, но в случае опасности они были способны перейти на бег на двух ногах (об этом говорит то, что примерно 80% веса тела приходилось на задние конечности).
Хотя этозавры были достаточно подвижными и неплохо защищенными травоядными, но при увеличении численности хищных динозавров и ухудшении условий внешней среды (вероятный рост содержания углекислого газа в атмосфере в конце триаса) они оказались неконкурентоспособны.
Состав Отряда (Семейства).
Как уже говорилось все представители Aetosauria входят в единственное семейство Stagonolepididae, в котором выделяют два подсемейства: Desmatosuchinae (крупные этозавры с защитными рогами на шее) включает роды Acaenasuchus, Desmatosuchus, Longosuchus, Lucasuchus и Sierritasuchus; и Typothoracisinae (крупные и широкие этозавры без рогов) включающий Typothorax, Redondasuchus, Tecovasuchus, Rioarribasuchus, Paratypothorax.
Остальных этозавров иногда объединяют в подсемейство Aetosaurinae, но его сейчас считают парафилетическим, поэтому роды Aetosaurus, Calyptosuchus, Coahomasuchus, Neoaetosauroides, Stagonolepis сейчас не сгруппированы.
|
Все, что нужно, можно найти прямо здесь: |
 |
| ad-place | ||
:-)
Группа сайтов от Юры(J) до Силура(S). Вы зашли в Триас.