
Завроптеригии триаса.
Характеристика группы.
Примерно в то время, когда первые ихтиоптеригии освоили океаны, начался переход к жизни в море другой группы рептилий, завроптеригий. Хотя перестройка их тела была менее глубокой, чем у ихтиоптеригий, она потребовала больше времени. И лишь в конце триаса зауроптеригии стали хозяевами океанов.
Завроптеригии – это надотряд Sauropterygia в подклассе чешуйчатых рептилий, хотя их ранг в мезозойской фауне повыше ранга надотряда. Завроптеригии были морскими рептилиями, которые смогли выработать особый стиль передвижения в виде. Этот стиль был похож на полет, причем некоторые зауроптеригии летали, используя только передние плавники, а другие – и передние, и задние. Из современных морских животных так передвигаются только морские черепахи, которые могут быть дальними родственниками завроптеригий. Завроптеригии все были морскими хищниками, питавшимися как малоподвижными морскими животными, так и подвижными рыбами, морскими рептилиями и даже птерозаврами.
Предполагается, что завроптеригии происходят от диапсидных предков, однако они, как и ихтиоптеригии, утратили нижние височные окна и имели эвриапсидный череп. Но если ихтиоптеригии имели череп по типу дельфина с коротким заглазничныи отделом и длинной мордой, то у завроптеригий заглазничная часть черепа была увеличена, а верхние окна удлинены.
Дышали завроптеригии, как и другие морские рептилии, через ноздри. Ноздри у них были немного смещены к глазам, однако при длинной и гибкой шее завроптеригиям не нужно было сильно смещать ноздри к глазницам. В остальном же череп завроптеригий напоминает черепа примитивных диапсидных рептилий. Это такой же слабый подвижный череп с весьма легкими челюстями, усаженными небольшими коническими зубами. (В юре появятся крупные завроптеригии с более мощными зубами.)
В ходе эволюции череп завроптеригий получил некоторые черты, отличающие его от наземных предков. Кроме утраты нижних височных окон, это утрата слезных, таблитчатых, заднетеменных и надвисочных костей. Также зауроптеригии обзавелисб длинным ретроартикулярным отростком нижней челюсти.
Важной отличительной особенностью зауроптеригий была удлиненная шея. И в отличие от современнных жирафов или триасовых проторозавров шея у них, удлинялась за счет увеличения количества шейных позвонков, их может быть до 20 штук. Туловищный отдел имеет примерно такую же длину и каждый туловищный позвонок соединен с ребрами. Что странно для ящеров с примитивным строением, у зауроптеригий в крестце 3-4 позвонка, которые им были совершенно не нужны при передвижении в воде. Ну а хвостовой отдел у завроптеригий постепенно сокращается по ходу эволюции, хотя у ранних представителей группы он длинный и приспособлен для плавания. Важная особенность позвоночника завроптеригий – отсутствие интерцентров в позвонках.
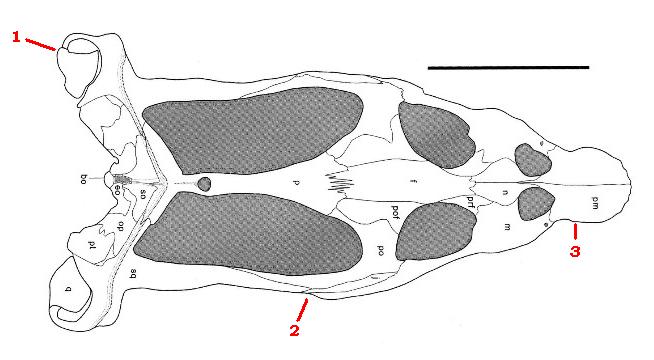
Рисунок, изображающий череп нотозавра (вид Nothosaurus yangjuanensis), вид сверху. Это типичный представитель триасовых зауроптериригий с их типичными отличительными чертами. Цифрами отмечены: 1) выступающий за границы черепа комплекс челюстного сустава; 2) конец зубного ряда верхнней челюсти далеко позади глазниц, а также (чуть выше) скуловая кость, ставшая узкой полоской; 3) ростр, сормированный предчелюстными костями, которые уходят далеко назад между ноздрей.
---
В ходе эволюции наиболее сильно были преобразованы пояса конечностей. У завроптеригий возникли плечевой и тазовый щиты, которые позволяли совершать мощные и быстрые гребковые движения вперед и вниз-назад. Передний щит был сформирован (в основном) сросшимися по средней линии груди коракоидами, лопатки также срастались впереди или соединялись срастающимися ключицами (лопатка была отделена от коракоида и налегала на ключицу), межключица редуцировалась до небольшой кости между ключицами или между лопатками. При этом вертикальный вырост лопатки, который был необходим на суше, также очень сильно редуцировался.
Задний щит состоял из разросшихся седалищных и лобковых костей, в тоже время подвздошные кости уменьшались в размере. Таким образом, контакт между позвоночником и поясами конечностей был почти потерян. А для передачи усилия конечностей всему телу завроптеригии были вынуждены увеличить и усилить брюшные ребра – гастралии. Они придавали прочность нижней стороне тела зауроптеригий и передавали тяговое усилие с конечностей на ребра.
Преобразование конечностей шло по пути сокращения костей предплечья и голени, а также по пути увеличения числа фаланг пальцев. Но при этом завроптеригии сохраняли длинные плечевые и бедренные кости. И их особенностью была изогнутая плечевая кость.
Направления эволюции завроптеригий в триасе.
В раннем и среднем триасе зауроптеригии были плохо адаптированы к жизни в воде. Их хвост был длинным, но слабым, а лапы еще не были преобразованы в плавники. Площадь плечевого и тазового щитов была мизерной. Скорее всего эти животные больше ползали по дну, чем плавали. Но для обитания в приливной зоне и на рифах этого было достаточно.
В дальнейшем эволюция шла по пути преобразования лап в плавники, сокращения хвоста и увеличения мощи плавательных мышц. При этом относительная длина шеи росла медленно, больше росла ее гибкость. Зато размер тела зауроптеригий рос очень быстро.
Одна из ветвей зауроптеригий (плиозавры) пошла по пути сокращения шеи и увеличения головы. В юрском периоде эти плиозавры стали крупнейшими морскими хищниками.
Подгруппы завроптеригий в триасе.
Триас – время примитивных завроптеригий. Они представлены несколькими примитивными образцами неясного систематического положения (условно поделены на Пахиплеврозавров (Pachypleurosauria) и Simosauridae), а также большой группой Нотозавры (Nothosauridae), вымершей в конце Триаса. Эти завроптеригии еще вели полуводный образ жизни, проводя значительную часть времени на берегу и, видимо, используя солнечное тепло для повышения температуры тела.
Группа, объединяющая более совершенных плиозавров и плезиозавров, возникла ближе к концу триаса и не была успешной до конца периода, так как ее представители пока еще не могли быстро передвигаться в воде.
|
Все, что нужно, можно найти прямо здесь: |
 |
| ad-place | ||
:-)
Группа сайтов от Юры(J) до Силура(S). Вы зашли в Триас.