
Ихтиоптеригии триаса.
Характеристика группы.
В раннем триасе состоядась первая удачная попытка рептилий вернуться в водную среду. В это время в океанах появились первые ихтиозавры (точнее ихтиоптеригии). За несколько миллионов лет они стали главенствующими океанскими хищниками, достигли гигантских размеров и стали очень разнообразны. Однако, век гигантских ихтиозавров оказался очень короток. Уже к раэтскому веку гиганты исчезают, а разнообразие остальных ихтиозавров резко снижается. Им на смену приходят другие морские рептилии – завроптеригии. Тем не менее, не смотря на все проблемы, ихтиоптеригии пережили вымирание в конце триаса и продолжили существование в юре.
Ихтиозавры – это отряд Ichthyosaurs или надотряд Ichthyopterygia (Ихтиоптеригии) класса рептилий. Считается, что ихтиоптеригии происходят от неизвестных пермских диапсидных рептилий, хотя они очень непохожи на своих предков. Все ихтиоптеригии были адаптированы для постоянной жизни в воде, считается, что ни один из известных представителей надотряда Ichthyopterygia не мог выходить на сушу. То есть все они проводили всю жизнь в воде. Самые примитивные обитали на мелководье: в мелких лагунах, у рифов, в дельтах рек. А более совершенные обитали в глубоводных бассейнах, постоянно курсируя между поверхностью (где они дышали) и большими глубинами (где они охотились).
Ихтиоптеригии являлись очень своеобразными рептилиями. Для них не только не известны переходные формы между наземными и водными видами; не удается даже определить близкородственные им формы диапсид, да и отнесение их к диапсидам весьма условно. Главное их отличие от других диапсидных ретилий – отсутствие нижних височных окон. (Это так называемый эуриапсидный тип черепа.) Отсутствие нижних окон, конечно, существенное отличие от диапсид, однако, оно не позволяет считать ихтиоптеригий особой ветвью рептилий. Дело в том, что закрытие височных окон является обычнм делом для рептилий, переходящих к жизни в воде.
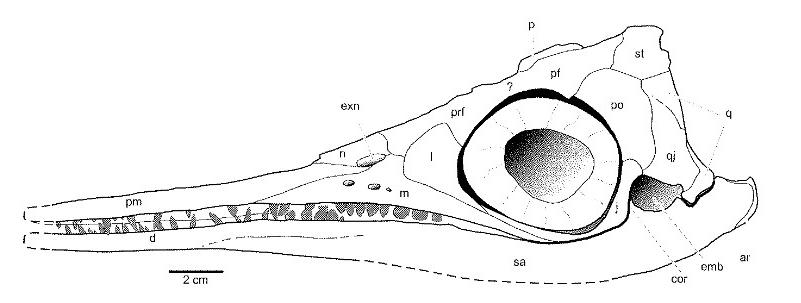
Рисунок, изображающий череп среднетриасового ихтиозавра Миксозавра (Mixosaurus) при взгляде сбоку. Обратите внимание на огромные глазницы, скуловую область с вырезкой и строение морды, в которой при адаптации к жизни в воде произошло смещение ноздрей назад, а предчелюсть вытянулась. К сожалению плохо виден теменной гребень.
---
Для ихтиозавров был характерен череп, напоминающий череп дельфина: длинное рыло, сформированное предчелюстными костями, слабые челюсти с мелкими коническими зубами (они имели текодонтное или субтекодонтное крепление), иногда зубы более крупные. Дыхание осуществлялось через крупные ноздри, смещенные назад к глазницам. У прогрессивных форм задненижний край ноздри формировался короткой слезной костью. Поэтому ноздря заканчивалась позади переднего края глазницы. Глазница очень большая, и судя по всему она вмещала не менее крупное глазное яблоко, которое могло достигать 20 см в диаметре, что в настоящее время характерно для морских животных больших глубин. Кроме того, для ихтиоптеригий характерны очень узкая скуловая кость и расположение челюстного сустава выше нижнего края глазницы. Поскольку ихтиозавры происходили от достаточно примитивных рептилий, у них сохранялось теменное отверстие в черепе.
Поскольку считается, что ихтиозавры имели конвергентное сходство с рыбами, то шейного перехвата у них не было. Однако, это не относится к ранним ихтиоптеригиям, у которых шея была хоть и короткой, но различимой. Более того, и в скелетах более поздних ихтиозавров шейный отдел различим. В туловищном отделе количество позвонков очень велико. 30-50 штук – нормальное дело для ихтиозавров. У некоторых, змеевидных, форм оно может быть значительно больше. Ко всем туловишным позвонкам присоединяются ребра. Длина хвостового отдела позвоночника сопоставима с длиной остального тела. У примитивных форм он длиннее, поскольку считается, что они плавали за счет волнообразных движений всего хвоста, как современные крокодилы или морские змеи. но в ходе эволюции хвост ихтиозавров стал короче, а на его конце сформировался хвостовой плавник. В отличие от всех других позвоночных, у ихтиозавров позвоночник продолжался в нижнюю лопасть плавника. А в том месте, где от хвоста отходила верхняя лопасть, позвоночник имел характерный двойной изгиб: сначала он изгибался вверх, а потом уходил вниз.
Конечности ихтиозавров были преобразованы в плавники и частично редуцированы. Каждый плавник состоял из сросшихся вместе пальцев, у которых произошло увеличение числа фаланг. Сверху лопасти плавника были покрыты жировой тканью, кожей и чешуей, что обеспецивало им прочность и обтекаемость. При этом ихтиозавры мало использовали плавники при плавании. Лишь некоторые виды активно использовали передние плавники. А задние имели гораздо меньшее значение, лишь играли роль дополнительных стабилизаторов. Поэтому они в ходе эволюции они были сильнее редуцированы, чем передние. Эволюция же передних конечностей шла в сторону поднятия их с нижней стороны тела ближе к его средней линии. Пояс передних конечностей при этом сильно уменьшился в размерах и распался на две не соединеные части. Тазовый пояс постепенно практически исчез, однако у самых ранних ихтиоптеригий он сохранялся. Два выраденных крестцовых позвонка соединялись удлиненными крестцовыми ребрами с небольшой подвздошной костьью, к которой были присоединены лобковая и седалищная, формировавшие горизонтальную пластину на нижней стороне тела. Такая конструкция таза говорит о том, что либо первые триасовые ихтиоптеригии могли выходить на сушу, либо их непосредственные предки сохраняли такую возможность.
Известно, что кожа ихтиозавров была лишена чешуи, возможно она постилалась жировой тканью, а сверху покрыта слизью. Из мягких тканей кожи и подкожных структур у ихтиозавров формировался спинной плавник, но он существовал не у всех видов.
В воде ихиозавры, в основном, полагались на зрение, об этом говорят их очень крупные глаза. Возможно, они смогли восстановить присущие рыбам и амфибиям органы боковой линии, чтобы ощущать колебания воды. В то же время слух у них был слаб, а электрорецепции и подводного обоняния не было. Т.е. на предельных глубинах в полной тепноте ихтиозавр не мог быть эффективным хищником.
Поскольку ихтиоптеригии не выходили на сушу, они дожны были перейти к живорождению. Считается, что это было не настоящее живорождение, а яйцеживорождение, когда зародыш развивался внутри яйца, не завися от матери.
Направления эволюции ихтиоптеригий в триасе.
В начале триаса ихтиоптеригии не были полностью адаптированы к жизни в океане. Поэтому первые из них просто приспосабливались к водной среде, теряя связь с сушей. У них улучшалась обтекаемость тела, увеличивался хвостовой плавник, сокращались конечности.
После появления настоящих ихтиозавров развтие пошло по двум направлениям. Одна ветвь привела к возникновению гигантов – доминирующих хищников океана. А другая ветвь привела к возникновению быстроходных и обладающих развитыми органами чувств юрских Офтальмозавров.
Подгруппы ихтиоптеригий в триасе.
Ранние ихтиоптеригии представлены малочисленными семейства некрупных ихтиозавров: Utatsusauridae, Parvinatatoridae, Гриппииды (Grippiidae), (возможно) Omphalosauridae.
А настоящие ихтиозавры в триасе представлены более примитивными семействами Миксозавридами (Mixosauridae), Цимбоспондилиды (Cymbospondylidae), Шастазавриды (Shastasauridae) и другими; а также более совершенными дельфиноподобными ихтиозаврами из групп Parvipelvia и Euichthyosauria (например, сем. Californosauridae).
|
Все, что нужно, можно найти прямо здесь: |
 |
| ad-place | ||
:-)
Группа сайтов от Юры(J) до Силура(S). Вы зашли в Триас.